| ||||
 | |||||
Die Navigation auf dieser Website funktioniert am besten mit aktiviertem Java-Script | |||||
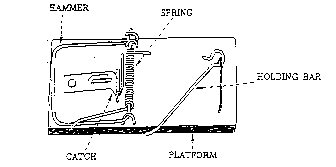
Molekulare Maschinen Experimentelle Bestätigung einer intelligenten Planung von Michael J. Behe Michael J. Behe ist Außerordentlicher Professor der Chemie an der Lehigh Universität in Pennsylvania und Partner des "Discovery Institute for Renewal of Science & Culture". Eine Serie von Augen Wie sehen wir? Im 19. Jahrhundert war die Anatomie des Auges detailliert bekannt und die ausgefeilten Mechanismen, die es einsetzt, um ein akkurates Bild der äußeren Welt zu liefern, erstaunte jeden, der mit ihnen vertraut war. Wissenschaftler des 19. Jahrhunderts beobachteten richtigerweise, daß, falls einer Person irgendeine der vielen integrierten Ausrüstungsmerkmale des Auges - wie die Linse, die Iris oder die okularen Muskeln – fehlte, das unausweichliche Ergebnis ein schwerer Verlust des Sehens oder gar totale Blindheit sein würde. So schloß man, daß das Auge nur dann funktionieren könne, wenn es fast ganz intakt sei. Da Charles Darwin mit möglichen Einwänden gegen seine Theorie der Evolution durch natürliche Auslese rechnete, besprach er das Problem des Auges in einem Kapitel seines Buches „Ursprung der Arten“, das den passenden Titel „Organe extremer Perfektion und Kompliziertheit“ trug. Er war sich bewußt darüber, daß, falls ein Organ von der Kompliziertheit des Auges in nur einer Generation plötzlich erschien, dieses Ereignis gleichbedeutend wäre mit einem Wunder. Irgendwie mußte die Schwierigkeit, welche die Öffentlichkeit mit der graduellen Bildung komplexer Organe hatte, ausgeräumt werden, sollte die Darwinsche Evolutionstheorie glaubwürdig erscheinen. Darwin tat dies brillant, nicht durch die tatsächliche Beschreibung eines Entwicklungswegs, den die Evolution benutzt haben könnte, um das Auge zu konstruieren, sondern eher durch die Hinweise auf eine Vielfalt von Tieren, von denen man wußte, daß sie Augen verschiedener Konstruktionsweisen hatten, die von einem einfachen lichtempfindlichen Punkt bis zur komplexen Kamera der Vertebraten reicht, und dadurch nahe legend, daß die Evolution des menschlichen Auges ähnliche Organe als Zwischenschritte hätte enthalten können. Aber die Frage bleibt: wie sehen wir? Obwohl Darwin die Mehrheit der Welt überreden konnte, daß ein modernes Auge graduell aus einer viel simpleren Struktur hervorgebracht werden könnte, hat er nicht einmal versucht, zu erklären, wie der einfache lichtempfindliche Punkt, der ja sein Ausgangspunkt war, tatsächlich arbeitet. In seiner Besprechung des Auges hat Darwin die Frage seines grundlegenden Mechanismus fallen gelassen.1 Wie ein Nerv dahin kommt, lichtempfindlich zu sein, berührt uns wohl kaum so sehr, wie die Frage, wie das Leben selbst entstand. Er hatte ausgezeichnete Gründe dafür, die Antwort auf diese Frage fallen zu lassen. Die Wissenschaft des 19. Jahrhunderts war noch nicht zu dem Punkt vorgedrungen, von dem aus man sich dieser Frage auch nur nähern konnte. Die Frage, wie das Auge arbeitet – d. h. was geschieht, wenn ein Lichtphoton erstmals auf die Retina trifft – konnte zu der Zeit einfach nicht beantwortet werden. Tatsächlich konnte keine Frage über den zugrunde liegenden Mechanismus des Lebens zu der Zeit beantwortet werden. Wie erzeugen Tiermuskeln Bewegung? Wie arbeitet die Photosynthese? Wie wird aus Nahrung Energie gezogen? Wie bekämpft der Körper eine Infektion? Niemand wußte das. „Calvinismus“ Nun, es scheint eine Charakteristik des menschlichen Geistes zu sein, daß er sich leicht simple Schritte vorstellen kann, die von Nicht-Funktion zu Funktion führen, wenn er nicht in das Wissen vom Mechanismus eines Prozesses gezwungen ist. Ein fröhliches Beispiel davon, kann man in dem populären Comic-Strip „Calvin and Hobbes“ betrachten. Der Steppke Calvin erlebt in der Gegenwart seines Tigers Hobbes immer wieder Abenteuer, indem sie in eine Kiste springen und Zeitreisen in die Vergangenheit machen oder wenn Calvin sich eine Spielzeugstrahlenpistole schnappt und sich selbst in verschiedene Tiergestalten verwandelt oder indem sie wiederum eine Kiste als Duplikator benutzen und von sich selbst Kopien herstellen, die es dann mit den Kräften der Welt in Gestalt seiner Mutter und seiner Lehrer aufnehmen. Für ein kleines Kind wie Calvin ist es leicht, sich vorzustellen, daß eine Kiste wie ein Flugzeug (oder sonstwas) fliegen kann, weil Calvin nicht weiß, wie ein Flugzeug funktioniert. Ein gutes Beispiel aus der biologischen Welt komplexer Veränderungen, die einfach erscheinen, ist der Glaube an spontane Erzeugung. Einer der prominentesten Vertreter der spontanen Erzeugung im mittleren neunzehnten Jahrhundert war Ernst Haeckel, ein großer Bewunderer Darwins und ein eifriger Popularisierer der Darwinschen Theorie. Aufgrund der begrenzten Ansicht von Zellen, welche die damals vorhandenen Mikroskope hergaben, glaubte Haeckel, daß die Zelle „einfache kleine Klumpen eiweißhaltiger Mischungen von Karbon“ seien,2 nicht viel anderes als ein Stück mikroskopische Götterspeise. Dadurch erschien es Haeckel so, als könnte solch simples Leben mit Leichtigkeit aus unbelebtem Material hergestellt worden sein. Im Jahre 1859, dem Erscheinungsjahr von „Ursprung der Arten“, holte ein Forschungsschiff, H.M.S. „Cyclops“, etwas merkwürdig aussehenden Schlamm vom Meeresboden herauf. Eines Tages kam Haeckel, um den Schlamm zu untersuchen, und dachte, es erinnere ihn sehr an einige Zellen, die er unter einem Mikroskop betrachtet hatte. Aufgeregt brachte er dies Thomas Henry Huxley zur Kenntnis, Darwins großem Freund und Verfechter. Huxley kam auch zu der Überzeugung, daß es „Urschleim“ (=Protoplasma) wäre, der Erzeuger des Lebens selbst und Huxley gab dem Schlamm den Namen „Bathybius Haeckelii“, nach dem berühmten Verfechter der Abiogenese. Der Schlamm wollte nicht wachsen. In späteren Jahren, mit der Entwicklung neuer biochemischer Techniken und verbesserten Mikroskopen, wurde die Komplexität der Zelle enthüllt. Die „einfachen Klumpen“ zeigten auf einmal Tausende verschiedenster Typen organischer Moleküle, nukleotide Säuren, viele separate subzellulare Strukturen, spezialisierte Abteilungen für spezielle Prozesse und eine äußerst komplizierte Architektur. Wenn wir aus der Perspektive unserer Zeit zurück blicken, erscheint uns die Episode mit dem „Bathybius Haeckelii“ albern oder geradezu peinlich, aber das sollte sie nicht. Haeckel und Huxley benahmen sich ganz natürlich, so wie Calvin: da sie die Komplexität von Zellen nicht kannten, war es leicht für sie, zu glauben, daß Zellen aus einfachem Schlamm stammten. In der Geschichte hat es viele andere solcher Beispiele gegeben, ähnlich dem von Haeckel, Huxley und der Zelle, in denen ein Kernstück wissenschaftlichen Puzzles jenseits des Verständnisses der betreffenden Zeit war. Es gibt in der Wissenschaft sogar eine wunderliche Bezeichnung für eine Maschine, Struktur oder einen Prozeß, die irgend etwas tun, deren Wirkungsweise jedoch unbekannt sind: so etwas nennt man eine „Black Box“. Zu Darwins Lebzeiten war die gesamte Biologie eine solche „Black Box“, nicht nur die Zelle, das Auge, die Verdauung oder das Immunsystem, sondern jede biologische Struktur und Funktion, weil letztlich niemand erklären konnte, wie biologische Prozesse vor sich gingen. Ernst Mayr, der prominente Biologe, Historiker und die treibende Kraft hinter der neo-darwinistischen Synthese, hat aufgezeigt, daß : 3 Jegliche wissenschaftliche Revolution alle möglichen Arten von „Black Boxes“ hin nehmen muß, weil man niemals irgendwelche begrifflichen Fortschritte machen würde, wenn man warten würde, bis alle „Black Boxes“ geöffnet worden seien. Das ist wahr. Aber in früheren Zeiten schien sich die Wissenschaft und mitunter sogar die ganze Welt zu verändern, wenn „Black Boxes“ geöffnet worden waren. Die Biologie hat sich aufgrund des Darwinschen Modells enorm weiter entwickelt. Aber die „Black Boxes“, die Darwin als ungeöffnet hin nahm, sind heute geöffnet worden und unsere Sicht der Welt wird erneut erschüttert. Proteine Um die molekulare Basis des Lebens zu verstehen, ist es nötig, zu verstehen, wie solche Dinge wie „Proteine“ arbeiten. Obwohl die meisten Menschen an Proteine als etwas Eßbares denken, als eine der Hauptnahrungsgruppen, dienen Proteine in ungegessenen Tieren und Pflanzen einem völlig anderen Zweck. Proteine bilden die Maschinerie lebenden Gewebes, das die Strukturen baut und die nötigen chemischen Reaktionen ausführt, die zum Leben notwendig sind. Zum Beispiel – der erste von vielen Schritten, die für die Umwandlung von Zucker zu biologisch brauchbaren Energieformen nötig sind, wird von einem Potein ausgeführt, das „Hexokinase“ genannt wird. Die Haut wird zum großen Teil von einem „Collagen“ genannten Protein gebaut. Wenn Licht auf Ihre Retina trifft, reagiert sie zuerst mit einem Protein, das man „Rhodopsin“ nennt. Wie man schon aus diesen wenigen begrenzten Beispielen ersehen kann, führen Proteine erstaunlich unterschiedliche Funktionen aus. Im allgemeinen jedoch kann ein bestimmtes Protein nur eine oder wenige Funktionen ausführen: Rhodopsin kann keine Haut bilden und Collagen kann nicht nutzbringend mit Licht reagieren. Deshalb enthält eine typische Zelle Tausende und Abertausende verschiedene Arten von Proteinen, um die vielen nötigen Prozesse des Lebens ausführen zu können, ganz ähnlich einer Tischlerwerkstatt, die viele verschiedene Arten von Werkzeug enthält, um allerlei Tischlerarbeiten ausführen zu können. Wie sehen diese vielseitigen Werkzeuge aus? Die grundlegende Struktur von Proteinen ist recht einfach: sie sind gebildet aus einer zusammenhängenden Kette separater Untereinheiten, den „Aminosäuren“. Obwohl die Proteinketten aus fünfzig bis eintausend Aminosäureverbindungen bestehen können, kann jede Stelle nur eine von zwanzig verschiedenen Aminosäuren enthalten. Sie ähneln auf diese Weise Wörtern: Wörter gibt es in verschiedenen Längen, aber sie sind aufgebaut aus einer Menge von 26 separaten Buchstaben. Nun schwimmen die Proteine nicht herum wie eine schlotterige Kette, sondern falten sich vielmehr zu einer sehr präzisen Struktur zusammen, die für verschiedene Typen von Proteinen sehr unterschiedlich sein kann. Letzen Endes können zwei unterschiedliche Aminosäuresequenzen – zwei verschiedene Proteine – zu Strukturen gefaltet werden, die so verschieden von einander sein können wie ein 12-Zoll Schraubenschlüssel und eine Laubsäge. Und genau so wie Haushaltswerkzeuge können auch Proteine ihre spezifischen Aufgaben nicht mehr durchführen, wenn ihre Gestalt beträchtlich verzerrt wird. Das Sehvermögen des Menschen Biologische Prozesse auf molekularer Ebene werden im allgemeinen von einem Netzwerk aus Proteinen durchgeführt, von denen jedes Glied eine besondere Aufgabe in einer Kette erledigt. Laßt uns zu der Frage zurück kehren: wie sehen wir? Obwohl für Darwin das ursprüngliche Ereignis der Sehkraft eine „Black Box“ war, ist eine Antwort auf die Frage des Sehens durch die Anstrengungen vieler Biochemiker in Sicht.4 Wenn Licht auf die Retina trifft, wird ein Photon von einem organischen Molekül namens „11-cis-retinal“ absorbiert (aufgenommen) und veranlaßt es dadurch, sich in Pikosekunden zu „Trans-Retinal“ umzuorganisieren. Die Veränderung in der Gestalt des „11-cis-retinal“ führt einen korrespondierenden Wechsel in der Gestalt des Proteins „Rhodopsin“ herbei, an welches es fest gebunden ist. Als Folge dieser Metamorphose des Proteins, verändert sich das Verhalten des Proteins in sehr besonderer Weise. Das veränderte Protein kann nun mit einem anderen Protein, dem „Transducin“ interagieren. Bevor „Transducin“ mit sich mit „Rhodopsin“ vereinigt, ist es fest an ein kleines organisches Molekül namens „GDP“ gebunden, aber wenn es sich an „Rhodopsin“ bindet, trennt sich das „GDP“ von dem „Transducin“ und ein Molekül namens „GTP“, das zwar eng mit dem „GDP“ verwandt, aber doch entscheidend verschieden von ihm ist, bindet sich an das „Transducin“. Der Austausch des „GTP“ mit „GDP“ in dem „Transducinrhodopsin“-Komplex ändert sein Verhalten. „GTP-Transducinrhodopsin“ bindet sich mit einem Protein namens „Phosphodiesterase“ zusammen, das in der inneren Membrane der Zelle angesiedelt ist. Wenn es durch „Rhodopsin“ und seinem Gefolge gebunden wird, erlangt die „Phosphodiesterase“ die Fähigkeit, ein Molekül namens „cGMP“ zu spalten. Anfänglich gibt es einen Haufen von „cGMP“-Molekülen in der Zelle, aber die Tätigkeit der „Phosphodiesterase“ senkt die Konzentration des „cGMP“. Die Aktivierung der „Phosphodiesterase“ kann man mit dem Herausziehen eines Stöpsels in einer Badewanne vergleichen, in welcher so die Höhe des Wasserspiegels verringert wird. Ein zweites Membran-Protein, das „cGMP“ bindet, das ein „Ionen-Kanal“ genannt wird, kann man sich wie eine spezielle Ausfahrt vorstellen, welche die Anzahl der Sodium-Ionen in der Zelle reguliert. Normalerweise erlaubt der Ionen-Kanal Sodium-Ionen in die Zelle einzufließen, während ein separates Protein sie wieder hinaus pumpt. Die duale Tätigkeit des Ionen-Kanals und des Pump-Proteins hält das Niveau der Sodium-Ionen in der Zelle innerhalb eines engen Wertebereichs. Wenn die Konzentration des „cGMP“ von seinem normalen Wert durch die Spaltung der „Phosphodiesterase“ vermindert wird, schließen sich viele Kanäle und dadurch wird eine verminderte zellulare Konzentration positiv geladener Sodium-Ionen ausgelöst. Dies wiederum verursacht ein Ungleichgewicht von Ladungen über die ganze Zellmembran hinweg, welches dann letztlich einen (elektrischen) Strom auslöst, der durch den optischen Nerv zum Gehirn geschickt wird: das Ergebnis, wenn es vom Gehirn „interpretiert“ wird, ist das, was wir sehen. Wenn die Biochemie des Sehvermögens begrenzt wäre auf die oben beschriebenen Reaktionen, würde die Zelle sehr schnell ihren Vorrat an „11-cis-Retinal“ und „cGMP“ abbauen und ebenso wäre ihr Vorrat an Sodium-Ionen bald ausgeschöpft. Es muß also ein System existieren, um die Signale zu begrenzen und die Zelle in ihren Ausgangszustand zurück zu versetzen; tatsächlich gibt es mehrere Systeme, die das bewerkstelligen. Im Dunkeln erlauben die Ionen-Kanäle normalerweise, zusätzlich zu den Sodium-Ionen, auch Calcium-Ionen in sie einzuschwemmen; Calcium wird von verschiedenen Proteinen zurück gepumpt, um eine ausgeglichene intrazelluläre Calciumkonzentration zu ermöglichen. Wenn jedoch das „cGMP“-Niveau fällt und dadurch die Ionen-Kanäle sich schließen und auch die Sodium-Konzentration abnimmt, wird auch die Calcium-Konzentration verringert. Das „Phosphodiesterase“-Enzym, welches „cGMP“ zerstört, wird bei einer geringen Calcium-Konzentration sehr verlangsamt. Zusätzlich beginnt ein Protein, genannt „Guanylate Cyclase“, „cGMP“ zu re-synthetisieren, wenn das Calcium-Niveau sich zu senken beginnt. In der Zwischenzeit, während all das vor sich geht, wird „Metarhodopsin II“ durch ein Enzym, „Rhodopsin Kinase“, chemisch verändert, welches eine Phosphatgruppe auf ihr Substrat plaziert. Das modifizierte „Rhodopsin“ wird dann durch ein Protein, „synchronisiertes Arrestin“, gebunden, welches das „Rhodopsin“ daran hindert, weiteres „Transducin“ zu aktivieren. Auf diese Weise enthält die Zelle Mechanismen, um das verstärkte Signal, das durch ein einzelnes Photon ausgelöst wurde, zu begrenzen. „Trans-Retinal“ fällt schließlich aus dem „Rhodopsin“-Molekül aus und muß zurück verwandelt werden in „11-Cis-Retinal“ und wiederum durch „Opsin“ gebunden werden, um „Rhodopsin“ für einen neuen Seh-Zyklus zu regenieren. Hierfür wird „Trans-Retinal“ zuerst chemisch modifiziert durch ein Enzym zu „Transretinol“, eine Form, die zwei Wasserstoffatome mehr enthält. Ein zweites Enzym isomeriert das Molekül zu „11-Cis-Retinol“. Schließlich entfernt ein drittes Enzym die vorher hinzugefügten Wasserstoffatome, um „11-Cis-Retinol“ zu bilden – und der Zyklus ist vollendet. Zur Erklärung des Lebens Obwohl viele Einzelheiten der Biochemie des Sehens hier nicht angeführt wurden, soll der hier gegebene Überblick dazu dienen, aufzuzeigen, was letztlich damit gemeint ist, wenn man davon spricht das Sehen zu „erklären“. Dies ist die Ebene von Erklärung, die biologische Wissenschaft im Visier haben muß. Um sagen zu können, daß etwas von einer Funktionsweise verstanden wurde, muß jeder relevante Schritt in dem Prozeß erläutert werden. Die relevanten Schritte in biologischen Prozessen geschehen letztlich auf molekularer Ebene, deshalb muß eine befriedigende Erklärung eines biologischen Phänomens, wie das Sehen, die Verdauung, oder das Immunsystem, eine molekulare Erläuterung beinhalten. Jetzt, da die „Black Box“ des Sehens geöffnet worden ist, reicht es zu einer „evolutionistischen Erklärung“ dieser Kraft nicht mehr aus, nur die anatomischen Strukturen des ganzen Auges einzubeziehen, wie Darwin es im 19. Jahrhundert getan hat und wie die meisten Populärwissenschaftler es noch heute tun. Anatomie ist ganz einfach irrelevant. Genauso ist es mit dem fossilen Datensatz. Es spielt einfach keine Rolle, ob oder ob nicht die fossilen Datensätze mit der Evolutionstheorie übereinstimmen, genau so wenig wie es in der Physik eine Rolle spielte, ob Newtons Theorie mit alltäglichen Erfahrungen übereinstimmt. Die fossilen Datensätze haben uns nichts zu sagen darüber, sagen wir mal, ob oder wie die Interaktionen von „11-Cis-Retinal“ mit „Rhodopsin“, „Transducin“ und „Phosphodiesterase“ sich Schritt für Schritt entwickelt haben könnten. Genau so wenig bedeuten Muster der Biographie oder Bevölkerungsgenetik oder die Erklärungen, welche die Evolutionstheorie zu rudimentären Organen oder dem Reichtum der Arten abgegeben hat. „Wie ein Nerv lichtempfindlich wird, geht uns weniger an, als der Ursprung des Lebens“, sagte Darwin im 19. Jahrhundert. Aber beide Phänomene haben das Interesse moderner Biochemie angelockt. Die Geschichte der langsamen Paralyse der Forschung auf dem Gebiet des Ursprungs des Lebens ist ganz interessant, aber sie zu erzählen, dafür ist hier nicht der Raum. Es mag zu sagen genügen, daß momentan das Gebiet der Forschung des Ursprungs des Lebens sich aufgelöst hat in eine Kakophonie miteinander in Konflikt stehender Modelle, jedes von ihnen nicht überzeugend, ernsthaft unvollständig und vollständig inkompatibel mit konkurrierenden Modellen. Privat werden die meisten der evolutionistischen Biologen zugeben, daß die Wissenschaft keine Erklärung hat für den Ursprung des Lebens. 5 Die Absicht dieses Artikels ist es, zu zeigen, daß die selben Probleme, welche die Ursprung-des-Lebens-Forschung bedrängen, ebenfalls die Bemühungen erschweren, die zeigen sollen, wie praktisch jedes komplexe biologische System entstand. Die Biochemie hat eine molekulare Welt enthüllt, die entschieden jeglicher Erklärung durch jene Theorie widersteht, welche lange Zeit auf der Ebene des ganzen Organismus angewandt wurde. Keine von Darwins „Black Boxes“ – weder der Ursprung des Lebens oder der Ursprung des Sehens noch anderer komplexer biochemischer Systeme – ist durch seine Theorie erklärt worden. Unglaubliche Komplexität In seinem Buch „Der Ursprung der Arten“ stellte Darwin fest: 6 „Falls bewiesen werden könnte, daß irgendein komplexes Organ existiert, das nicht notwendigerweise durch zahlreiche, auf einander folgende, leichte Veränderungen geformt worden ist, dann würde meine Theorie total zusammen brechen.“ Ein System, das Darwins Kriterium entspricht, ist eines, das nicht reduzierbare Komplexität aufweist. Mit „nicht reduzierbarer Komplexität“ meine ich ein einzelnes System, das aus mehreren miteinander interagierenden Teilen besteht, welche zur grundlegenden Funktionalität beitragen, und in dem die Entfernung auch nur eines dieser Teile das gesamte System lahmlegt (das System funktioniert dann nicht mehr). Ein aus nicht reduzierbarer Komplexität bestehendes System kann nicht graduell (allmählich) aus leichten, auf einander folgenden Veränderungen eines Vorgängersystems gebaut werden, da jedes Vorgängersystem eines aus nicht reduzierbarer Komplexität bestehenden Systems per Definition nicht funktional ist (nicht funktionieren würde). Da natürliche Auslese eine Funktion zur Auslese benötigt, müßte ein aus nicht reduzierbarer Komplexität bestehendes biologisches System, falls es so etwas gibt, als eine integrierte Einheit zur natürlichen Auslese auftauchen und als solche irgend etwas haben, an dem sie arbeiten könnte. Es wird im allgemeinen fast überall anerkannt, daß ein solches plötzliches Ereignis unvereinbar wäre mit dem von Darwin vorgestellten Gradualismus. An diesem Punkt ist „nicht reduzierbar komplex“ jedoch nur ein Begriff, dessen Wirksamkeit hauptsächlich in seiner Definition liegt. Wir müssen uns nun fragen, ob es irgend etwas real existierendes gibt, welches tatsächlich nicht reduzierbar komplex ist, und, falls es so etwas gibt, sind irgendwelche nicht reduzierbaren Dinge auch biologische Systeme. Betrachten wir die einfache Mausefalle (Abb.1). Die Mausefalle, die meine Familie dazu benutzte, mit unerwünschten Nagern fertig zu werden, bestand aus einer Reihe von Teilen. Diese sind: (1) eine flache Holzplattform, die als Basis dient; (2) ein Metallhammer, der die eigentliche Arbeit tut, indem er die kleine Maus zerquetscht; (3) eine Drahtfeder mit verlängerten Enden, um gegen die Plattform und den Hammer zu drücken, wenn die Falle ausgelöst wird; (4) einen empfindlichen Schnapper, der nachgibt, wenn leichter Druck ausgeübt wird und (5) einen metallenen Querriegel, der den Hammer zurück hält, wenn die Falle geladen wird und mit dem Schnapper verbunden ist. Dann gibt es noch verschiedene Krampen und Schrauben, die das System zusammen halten.
Abbildung 1. Eine haushaltsübliche Mausefalle. Die mit einander arbeitenden Teile sind markiert. Falls eines der Teile fehlt, funktioniert die Mausefalle nicht. Falls irgend eines der Komponenten der Mausefalle (die Basis, der Hammer, die Feder, der Schnapper oder der Querriegel) entfernt wird, dann funktioniert die Mausefalle nicht mehr. Mit anderen Worten: die einfache, kleine Mausefalle ist erst dann fähig, eine Maus zu fangen, wenn alle einzelnen Teile zusammen gefügt worden sind. Weil die Mausefalle notwendigerweise aus mehreren Teilen besteht, ist sie nicht reduzierbar komplex. Folglich existieren nicht reduzierbar komplexe Systeme. Molekulare Maschinen Gibt es nun biochemische Systeme, die nicht reduzierbar komplex sind? Ja, es stellt sich heraus, daß viele von ihnen so sind. Oben haben wir Proteine besprochen. In vielen biologischen Strukturen sind Proteine einfach Komponenten (Bestandteile) größerer molekularer Maschinen. So wie ein Fernsehgerät aus Bildröhre, Drähten, Metallbolzen und Schrauben besteht, sind viele Proteine Teile von Strukturen, die nur funktionieren, wenn praktisch alle ihrer Komponenten zusammen gefügt worden sind. Ein gutes Beispiel dafür ist ein „Cilium“. 7 „Cilium“ Animation
Abbildung 2a Animation eines „Ciliums“
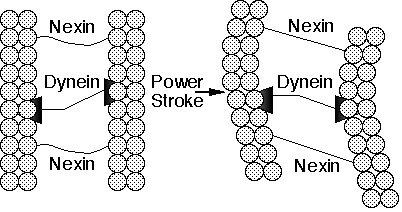
„Cilia“ sind haarähnliche Organellen auf der Oberfläche vieler tierischer und pflanzlicher Zellen, die dazu dienen, Flüssigkeit über die Oberfläche zu bewegen oder um einzelne Zellen durch eine Flüssigkeit zu „rudern“. Im Menschen, zum Beispiel, haben Saumepithelzellen, die den Belag des Atmungstraktes bilden, ungefähr 200 Cilias, die im Synchrontakt schlagen, um die Schleimschicht zur Beseitigung zur Gurgel hin zu treiben. Ein Cilium besteht aus einem membranbedeckten Bündel von Fibern, „Axoneme“ genannt. Ein Axonem enthält einen Ring aus 9 Doppel-Mikrotubuli, die zwei zentrale Mikrotubuli umringen. Jedes äußere Doppel besteht aus einem Ring aus 13 Filamenten (“Subfiber A“), verwachsen mit einer Baugruppe von 10 Filamenten („Subfiber B“). Die Filamente der Mikrotubuli bestehen aus zwei Proteinen, genannt „Alpha“- und „BetaTubulin“. Die 11 Mikrotubuli, die ein Axonem bilden, werden durch drei Arten von Verbindern zusammen gehalten: die „Subfibern A“ sind mit den zentralen Mikrotubuli verbunden durch radiale Speichen; angrenzende äußere Doppel sind durch Verknüpfer verbunden, die aus hoch elastischem Protein, „Nexin“, bestehen; und die zentralen Mikrotubuli sind verbunden durch eine Verbindungsbrücke. Schließlich trägt jede „Subfiber A“ zwei Arme, einen inneren und einen äußeren Arm, die beide das Protein „Dynein“ enthalten. Aber wie arbeitet nun ein Cilium? Experimente haben gezeigt, daß die ciliarische Bewegung geschieht durch die chemisch angetriebene „Wanderung“ des „Dynein“-Arms über einen Mikrotubulus und die benachbarte „Subfiber B“ eines zweiten Mikrotubulus hinauf , so daß die zwei Mikrotubuli aneinander vorüber gleiten (Abb. 2a und 2b). Die Protein-Querverbindungen zwischen Mikrotubuli in einem intakten Cilium hindern jedoch die benachbarten Mikrotubuli daran, weiter als eine kurze Strecke aneinander vorbei zu gleiten. Diese Querverbindungen konvertieren deshalb die dynein-induzierte Gleitbewegung in eine biegende, beugende Bewegung der gesamten Axoneme.
Abbildung 2b. Schematische Zeichnung eines Ciliums. Der Arbeitstakt (Power Stroke) des Motor-Proteins, „Dynein“, an einen Mikrotubulus angefügt, hin zu der „Subfiber B“ eines benachbarten Mikrotubulus bringt die Fibern dazu, aneinander vorüber zu gleiten. Das flexible Verbindungs-Protein, „Nexin“, wandelt die schiebende Bewegung in eine biegende, beugende Bewegung um. Nun lassen Sie uns zurück lehnen, die Arbeitsweise des Ciliums betrachten und überlegen, was es impliziert. Cilia bestehen aus mindestens einem halben Dutzend Proteinen: „Alpha-Tubulin“, „Beta-Tubulin“, „Dynein“, „Nexin“, Speichen-Protein und einem zentralen Brücken-Protein. Diese verknüpfen, vereinigen und koppeln sich, um eine Aufgabe durchzuführen, die ciliarische Bewegung, und alle diese Proteine müssen anwesend sein, damit das Cilium funktionieren kann. Falls die Tubuline abwesend sind, dann gibt es keine Filamente zum Gleiten; falls das „Dynein“ abwesend ist, dann bleibt das Cilium starr und unbeweglich; falls „Nexin“ oder die anderen verbindenden Proteine fehlen, dann fallen die Axoneme auseinander, wenn die Filamente gleiten. Was wir an dem Cilium also sehen, ist nicht nur profunde Komplexität, sondern auch nicht reduzierbare Komplexität in molekularem Maßstab. Erinnern wir uns daran, daß wir mit dem Begriff „nicht reduzierbare Komplexität“ einen Apparat meinen, der verschiedene, unterschiedliche Komponenten braucht, um als Ganzes funktionieren zu können. Meine Mausefalle muß eine Basis, einen Hammer, eine Feder, einen Schnapper und einen Querriegel haben, die alle zusammen arbeiten, haben, damit sie funktioniert. Ganz ähnlich muß ein Cilium, so wie es zusammen gestellt ist, die gleitenden Filamente, verbindende Proteine und Motor-Proteine haben, um sein Funktionieren sicher zu stellen. Bei Abwesenheit auch nur eines Teils dieser Komponenten wird der Apparat nutzlos. Die Komponenten einer Cilia sind einzelne Moleküle. Dies bedeutet, daß es keine „Black Boxes“ mehr gibt, die Komplexität des Ciliums ist endgültig, fundamental. Und ebenso wie die Wissenschaftler, als sie die Komplexität der Zelle kennen lernten, begriffen, wie dumm es war, zu glauben, das Leben entstehe in einem einzigen oder mehreren Schritten ganz einfach aus dem Meeres-Schlamm, so begreifen auch wir heute, daß das komplexe Cilium nicht einfach aus einem einzelnen oder mehreren Schritten „erreicht“ werden kann. Aber da die Komplexität des Ciliums nicht reduzierbar ist, kann es keinerlei funktionelle Vorgänger gehabt haben. Da das nicht reduzierbar komplexe Cilium keine funktionale Vorgänger gehabt haben kann, kann es nicht durch natürliche Auslese hervor gebracht worden sein, welche ein Kontinuum an Funktionalität benötigt, um zu funktionieren. Natürliche Auslese ist wirkungslos, wenn es keine Funktion gibt, die man wählen könnte. Wir können noch weiter gehen und sagen, daß, falls das Cilium nicht durch natürliche Auslese hervor gebracht werden kann, dann wurde das Cilium entworfen und geplant. Das Studium der „molekularen Evolution“ Andere Beispiele nicht reduzierbarer Komplexität sind im Überfluß vorhanden, einschließlich Aspekte von Protein-Transport, Blutgerinnung, gesperrter kreisförmiger DNA, Elektronen-Transport, das bakterielle Flagellum, Telomere, Photosynthese, Transkriptions-Steuerung und viele mehr. Beispiele nicht reduzierbarer Komplexität können praktisch auf jeder Seite eines biochemischen Fachbuchs gefunden werden. Aber wenn diese Dinge durch die Darwinsche Evolution nicht erklärt werden können, wie hat die wissenschaftliche Gemeinschaft diese Phänomene in den letzten vierzig Jahren betrachtet? Eine gute Stelle, um eine Antwort auf diese Frage zu bekommen, ist das „Journal of Molecular Evolution“ (im folgenden kurz JME genannt). JME ist ein Journal, das mit dem speziellen Ziel gegründet wurde, sich mit dem Thema zu beschäftigen, wie Evolution auf molekularer Basis statt findet. Es hat hohe wissenschaftliche Qualitätsanforderungen und wird von einer prominenten Gestalt auf diesem Gebiet heraus gegeben. In einer kürzlich erschienenen Ausgabe des JME wurden elf Artikel veröffentlicht, von denen alle elf sich einfach nur mit der Analyse von Protein oder DNA Sequenzen beschäftigten. Keiner der Artikel besprach detaillierte Modelle zu Zwischenstufen in der Entwicklung komplexer biochemischer Strukturen. In den letzten zehn Jahren hat JME 886 Artikel veröffentlicht. Von diesen besprachen 95 die chemische Synthese von Molekülen, von denen man annimmt, sie seien für den Ursprung des Lebens nötig, 44 schlugen mathematische Modelle vor, um die Sequenzanalyse zu verbessern, 20 bezogen sich auf die evolutionären Implikationen gegenwärtiger Strukturen, und 719 waren Analysen von Proteinen oder polynukleotider Sequenzen. Es gab nicht einen Artikel, der detaillierte Modelle für die Zwischenstufen in der Entwicklung komplexer biochemischer Strukturen besprach – weder in „Proceedings of the National Academy of Science“, „Nature“, „Science“, „Journal of Molecular Biology“ noch, meines Wissens, überhaupt in irgendeinem anderen Journal. Die Literatur über molekulare Evolution wird überwältigend dominiert von Sequenz-Vergleichen. Aber Sequenz-Vergleiche können die Entwicklung komplexer biochemischer Systeme nicht besser erklären, als Darwins Vergleiche einfacher und komplexer Augen ihm erklären konnten, wie das Sehen funktioniert. Auf diesem Gebiet ist die Wissenschaft verstummt. Das heißt, wenn wir folgern, daß komplexe biochemische Systeme entworfen und geplant („designed“) worden sind, widersprechen wir keinerlei experimentellen Resultaten und stehen in keinem Konflikt mit irgendwelchen theoretischen Studien. Experimente müssen nicht hinterfragt werden, aber die Interpretation aller Experimente muß heute nachgeprüft werden, genau so wie die Ergebnisse von Experimenten, die in Einklang mit einer Newtonschen Sicht des Universums in Einklang standen, neu interpretiert werden mußten, als die Wellen-Partikel-Dualität von Materie erkannt wurde. Schlußfolgerung Es wird oft gesagt, daß die Wissenschaft jedwede Folgerungen vermeiden sollte, die einen Geschmack des Übernatürlichen haben. Aber es scheint mir beides zu sein, schlechte Logik und schlechte Wissenschaft. Die Wissenschaft ist kein Spiel, in dem willkürliche Regeln dafür benutzt werden, zu entscheiden, welche Erklärungen erlaubt sind. Sie ist eher das Bemühen, wahre Feststellungen über die physikalische Realität zu treffen. Es ist gerade einmal sechzig Jahre her, da der Ausdehnungscharakter des Universums zuerst beobachtet wurde. Diese Tatsache suggerierte sofort ein seltsames Ereignis – daß irgendwann in der Vergangenheit, das Universum aus einer extrem kleinen Baugröße zu expandieren begann. Vielen Leuten erschien diese Folgerung beladen mit Untertönen eines übernatürlichen Ereignisses – die Schöpfung, der Anfang des Universums. Der prominente Physiker A. S. Eddington sprach wahrscheinlich für viele Physiker, indem er seinen Abscheu solch einer Feststellung gegenüber aussprach.8 Philosophisch betrachtet, erscheint die Feststellung eines zusammenhanglosen Anfangs der gegenwärtigen Ordnung der Natur mir abstoßend, so wie es für die meisten sein muß, denke ich; und selbst jene, die einen Beweis für den Eingriff eines Schöpfers begrüßen würden, werden wahrscheinlich überlegen, daß ein einzelnes Heraufentwickeln in irgendeiner entfernten Epoche nicht wirklich die Art von Beziehung zwischen Gott und Seiner Welt ist, die dem Geist befriedigend erscheint. Nichtsdestoweniger wurde die Hypothese des „Big Bang“ von Physikern bereitwillig übernommen und hat sich als ein sehr fruchtbares Paradigma über die Jahre hinweg erwiesen. Das, was ich sagen will, ist, daß die Physiker den Daten dahin folgten, wohin sie führten, obwohl einige dachten, dies Modell gäbe Hilfestellung und Trost für die Religion. Heutzutage, da die Biochemie Beispiele phantastisch komplexer Molekularsysteme vermehrt, Systeme, die selbst einem Versuch, zu erklären, wie sie denn entstanden sein könnten, abschreckend gegenüber stehen, sollten wir eine Lektion von den Physikern lernen. Die Folgerung der Planung und des Entwurfs fließt ganz natürlich aus den Daten hervor. Wir sollten nicht davor zurück schrecken. Wir sollten sie bereitwillig annehmen und darauf aufbauen. Zusammenfassend ist es wichtig, zu begreifen, daß wir „Design“ (intelligenter Entwurf und Planung, Anm. d. Übersetzers) nicht aus dem heraus folgern, das wir nicht wissen, sondern aus dem, das wir wissen. Wir folgern „Design“ nicht, um eine „Black Box“ (Unwissenheit, Anm. d. Übersetzers) zu erklären, sondern eine „Open Box“ (Wissen, Anm. des Übersetzers). Ein Mann aus einer primitiven Kultur, der ein Automobil sieht, könnte erraten, daß es durch den Wind oder durch eine unter der Motorhaube versteckte Antilope angetrieben sein könnte, aber wenn er die Motorhaube öffnet und die Maschine sieht, wird er sofort begreifen, daß sie „designed“ (durch eine Intelligenz entworfen und geplant, Anm. d. Übersetzers) wurde. In gleicher Weise hat die Biochemie die Zelle geöffnet, um zu untersuchen, was sie funktionieren läßt, und wir sehen, daß auch sie „designed“ wurde. Es war ein Schock für die Leute des 19. Jahrhunderts, als sie aus den Beobachtungen, welche die Wissenschaft gemacht hatte, entdeckten, daß viele Charakteristika der biologischen Welt durch das elegante Prinzip natürlicher Auslese erklärt werden konnten. Es ist ein Schock für uns, im 20. Jahrhundert, aus Beobachtungen, welche die Wissenschaft gemacht hat, zu entdecken, daß der fundamentale Mechanismus des Lebens nicht durch natürliche Auslese beschrieben werden kann, und deshalb „designed“ (durch eine Intelligenz entworfen und geplant, Anm. d. Übersetzers) wurde. Aber wir müssen mit unserem Schock so gut wir können fertig werden. Die Theorie der ungeleiteten Evolution ist tot, aber die Arbeit der Wissenschaft geht weiter. Dieser Artikel wurde ursprünglich im Sommer 1994 beim Treffen der „C. S. Lewis Society“ in der Cambridge Universität präsentiert. Verweise 1. Darwin, Charles (1872) Origin of Species 6th ed (1988), p.151, New York University Press, New York 2. Farley, John (1979) The Spontaneous Generation Controversy from Descartes to Oparin, 2nd ed, p.73, The Johns Hopkins University Press, Baltimore 3. Mayr, Ernst (1991) One Long Argument, p. 146, Harvard University Press, Cambridge 4. Devlin, Thomas M. (1992) Textbook of Biochemistry, pp.938954, WileyLiss, New York 5. Der Schönredner von der Washington Universität John Angus Campbell hat beobachtet, daß „riesige Gedankengebäude wie der Positivismus niemals wirklich sterben. Denkende Menschen verwerfen sie allmählich und machen sie unter sich sogar lächerlich, behalten aber die überzeugendsten brauchbaren Teile, um die Uninformierten abzuschrecken.“ Aus "The Comic Frame and the Rhetoric of Science: Epistemology and Ethics in Darwin's Origin," Rhetoric Society Quarterly 24, pp.2750 (1994). Dies stimmt sicherlich für die Art und Weise, wie die wissenschaftliche Gemeinschaft die Fragen über den Ursprung des Lebens behandelt. 6. Darwin, p.154 7. Voet, D. & Voet, J.G. (1990) Biochemistry, pp.11321139, John Wiley & Sons, New York 8. Cited in Jaki, Stanley L. (1980) Cosmos and Creator, pp.56, Gateway Editions, Chicago Copyright ©1997 Michael Behe. All rights reserved. International copyright secured. Übersetzt von Roland R. Noetzelmann 2006 aus dem Amerikanischen Ich danke ARN (Access Research Network) für ihre freundliche Genehmigung. ARN – findet man hier: http://www.arn.org/ Das Original-Dokument findet man hier: http://www.arn.org/docs/behe/mb_mm92496.htm
| |||||